一、细胞分裂的类型细胞分裂(cell division)可分为无丝分裂(amitosis)、有丝分裂(mitosis)和减数分裂(meiosis)三种类型。 无丝分裂又称为直接分裂,由R. Remark(1841)首次发现于鸡胚血细胞。表现为细胞核伸长,从中部缢缩,然后细胞质分裂,其间不涉及纺锤体形成及染色体变化,故称为无丝分裂。无丝分裂不仅发现于原核生物,同时也发现于高等动植物,如植物的胚乳细胞、动物的胎膜,间充组织及肌肉细胞等等。 有丝分裂,又称为间接分裂,由W. Fleming (1882)年首次发现于动物及E. Strasburger(1880)年发现于植物。特点是有纺锤体染色体出现,子染色体被平均分配到子细胞,这种分裂方式普遍见于高等动植物。 减数分裂是指染色体复制一次而细胞连续分裂两次的分裂方式,是高等动植物配子体形成的分裂方式。 二、有丝分裂有丝分裂过程是一个连续的过程,为了便于描述人为的划分为六个时期:间期(interphase)、前期(prophase)、前中期(premetaphase)、中期(metaphase)、后期(anaphase)和末期(telophase)。其中间期包括G1期、S期和G2期,主要进行DNA复制等准备工作。 (一)前期前期(图13-3)的主要事件是:①染色质凝缩,②分裂极确立与纺锤体开始形成,③核仁解体,④核膜消失。 前期最显著的特征是染色质通过螺旋化和折叠,变短变粗,形成光学显微镜下可以分辨的染色体,每条染色体包含2个染色单体。
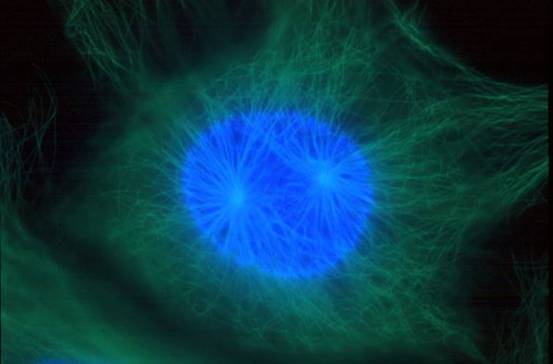
图13-3 前期两个中心体向两极移动(图片来自)
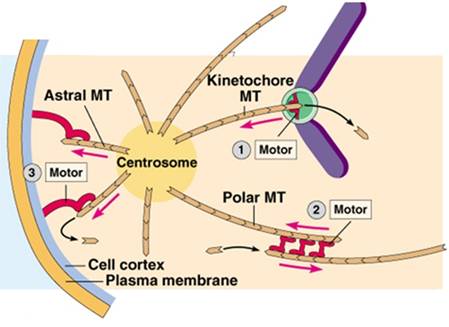
早在S期两个中心粒已完成复制,在前期移向两极,两对中心粒之间形成纺锤体微管,当核膜解体时,两对中心粒已到达两极,并在两者之间形成纺锤体,纺锤体微管包括 (图13-8): ①着丝点微管(kinetochore mt):由中心体发出,连接在着丝点上,负责将染色体牵引到纺锤体上,着丝点上具有马达蛋白。 ②星体微管(astral mt):由中心体向外放射出,末端结合有分子马达,负责两极的分离,同时确定纺锤体纵轴的方向。 ③极体微管(polar mt或overlap mt):由中心体发出,在纺锤体中部重叠,重叠部位结合有分子马达,负责将两极推开。
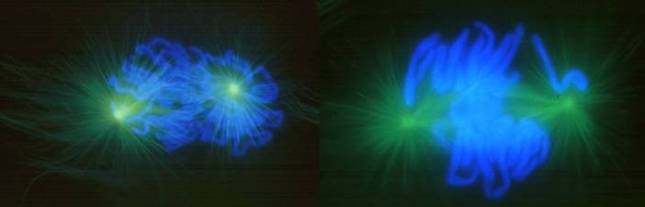
有两类马达蛋白参与染色体、分裂极的分离,一类是dynein,另一类是kinesin。 植物没有中心粒和星体,其纺锤体叫作无星纺锤体,分裂极的确定机理尚不明确。 (二)前中期指由核膜解体到染色体排列到赤道面(equatorial plane)这一阶段(图13-4)。纺锤体微管向细胞内部侵入,与染色体的着丝点结合。着丝点处的分子马达使染色体向微管的负端移动。在光镜下可以看到,此时染色体也就是既向一极移动也向另一极移动,是以振荡的方式移向纺锤体中部的。其原因是姊妹染色单体的着丝点都结合有微管和分子马达。
图13-4 左,前中期; 右,中期(图片来自) (三)中期指从染色体排列到赤道面上(图13-4右、13-5),到姊妹染色单体开始分向两极的一段时间,纵向观动物染色体呈辐射状排列。染色体两边的牵引力就像拔河一样达到平衡。
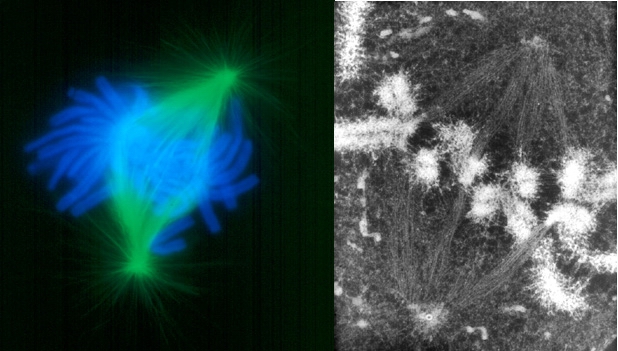
图13-5 中期,右图显示与染色体联接的微管(图片来自) (四)后期指姊妹染色单体分开并移向两极的时期,当子染色体到达两极后,标志这一时期结束(图13-6)。
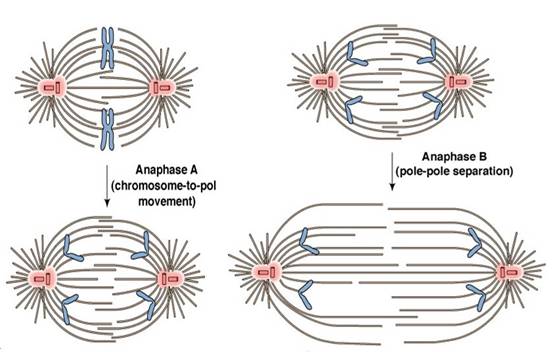
图13-6 后期姊妹染色单体分离(图片来自) 后期可以分为两个方面(图13-7):①后期A,指染色体向两极移动的过程。这是因为染色体着丝点微管在着丝点处去组装而缩短,在分子马达的作用下染色体向两极移动,体外实验证明即使在不存在ATP的情况下,染色体着丝点也有连接到正在去组装的微管上的能力,使染色体发生移动。②后期B,指两极间距离拉大的过程。这是因为一方面极体微管延长,结合在极体微管重叠部分的马达蛋白提供动力,推动两极分离,另一方面星体微管去组装而缩短,结合在星体微管正极的马达蛋白牵引两极距离加大。可见染色体的分离是在微管与分子马达的共同作用下实现的(图13-8)。 后期A,B是用xx鉴定出来的,如紫杉酚(taxol)能结合在微管的(+)端,抑制微管(+)端去组装,从而抑制后期A。动物中通常先发生后期A,再后期B,但也有些只发生后期A,还有的后期A、B同时发生。植物细胞没有后期B。
图13-7 后其A染色体分离,后期B两极延伸
图13-8 马达蛋白和微管系统共同协作,使染色体分离 (五)末期末期(图13-9)是从子染色体到达两极,至形成两个新细胞为止的时期。末期涉及子核的形成和胞质分裂两个方面。
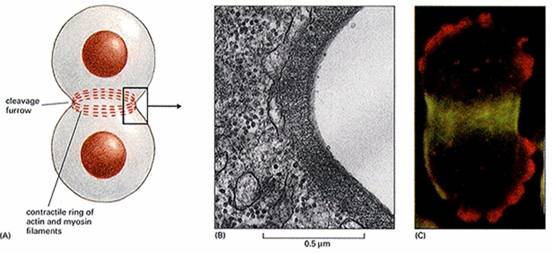
图13-9 末期 1、子核的形成末期子核的形成,大体经历了与前期相反的过程,即染色体解聚缩,核仁出现和核膜重新形成。核仁由染色体上的核仁组织中心形成(NORs),几个NORS共同组成一个大的核仁,因此核仁的数目通常比NORs的数目要少。 前期核膜解体后,核纤层蛋白B与核膜残余小泡结合,末期核纤层蛋白B去磷酸化,介导核膜的重新装配。 2、胞质分裂虽然核分裂与胞质分裂(cytokinesis)是相继发生的,但属于两个分离的过程,例如大多数昆虫的卵,核可进行多次分裂而无胞质分裂,某些藻类的多核细胞可长达数尺,以后胞质才分裂形成单核细胞。 动物细胞的胞质分裂是以形成收缩环的方式完成的(图13-10),收缩环在后期形成,由大量平行排列的肌动蛋白和结合在上面的myosin II等成分组成,用细胞松驰素及肌动蛋白和肌球蛋白抗体处理均能抑制收缩环的形成。不难想象胞质收缩环工作原理和肌肉收缩时一样的。
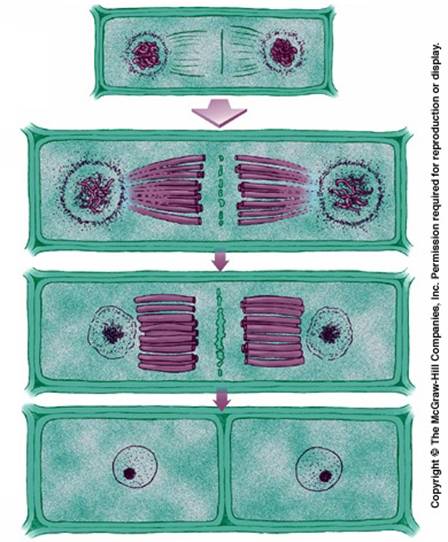
图13-10 动细胞的胞质收缩环 动物胞质分裂的另一特点是形成中体。末期纺锤体开始瓦解消失,但在纺锤体的中部微管数量增加,其中掺杂有高电子密度物质和囊状物,这一结构称为中体。在胞质分裂中的作用尚不清楚。 植物胞质分裂的机制不同于动物,后期或末期两极处微管消失,中间微管保留,并数量增加,形成桶状的成膜体(phragmoplast)。来自于高尔基体的囊泡沿微管转运到成膜体中间,融合形成细胞板。囊泡内的物质沉积为初生壁和中胶层,囊泡膜形成新的质膜,由于两侧质膜来源于共同的囊泡,因而膜间有许多连通的管道,形成胞间连丝。源源不断运送来的囊泡向细胞板融合,使细胞板扩展,形成完整的细胞壁,将子细胞一分为二(图13-11)。
图13-11 植物细胞成膜体的形成 |